
休斯顿动物园游记
在挣扎着写了一个月代码后,今天去动物园放放风,水篇博客找找灵感。先从比较众所周知的写起吧。
长颈鹿与喉返神经
这算是进化论里的经典案例了。喉返神经,顾名思义,一是用来控制喉部的,二是它走了弯路。虽然目的地是喉部,但它从大脑出来后却直奔心脏,绕过主动脉才又返回喉部。 这在人身上不是什么大问题,多走了几分米而已。但在长颈鹿身上,这条神经就多绕了四米多。神经信号一来一回花都谢了。有人认为这是长颈鹿很安静的原因,想说话嘴跟不上啊,想想都捉急。。但这还不是最惨的,已经灭绝的蜥脚类恐龙(就是《侏罗纪公园》里的那些长脖子),它们的喉返神经得多走上二三十米。这样的两只恐龙想对话大概就像拨号上网年代的网络聊天吧_(:з」∠)_
那进化是怎么把这么低效的设计保留下来的呢?这得从我们还在海里的时候讲起。
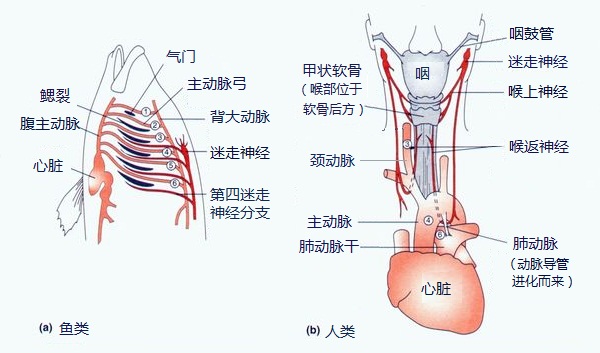
这个例子因为很好的体现了进化的短视性而被反复引述。进化不会把神经突然断开然后到另一边重新接起来,只会在第六动脉弓不断下移时,一点点地去延长喉返神经。"Inside Nature's Giants" 有一期解剖长颈鹿,专门拍摄了关于喉返神经的解剖,还非常合适的由道金斯在边上主持。(这视频在 YouTube 上,我在墙内实在没找到,试图自己上传结果优酷跟我说含有低俗内容(╯‵□′)╯︵┻━┻,所以想看的自己翻墙吧)更详细的讨论参考 Why Evolution Is True 的这部分。
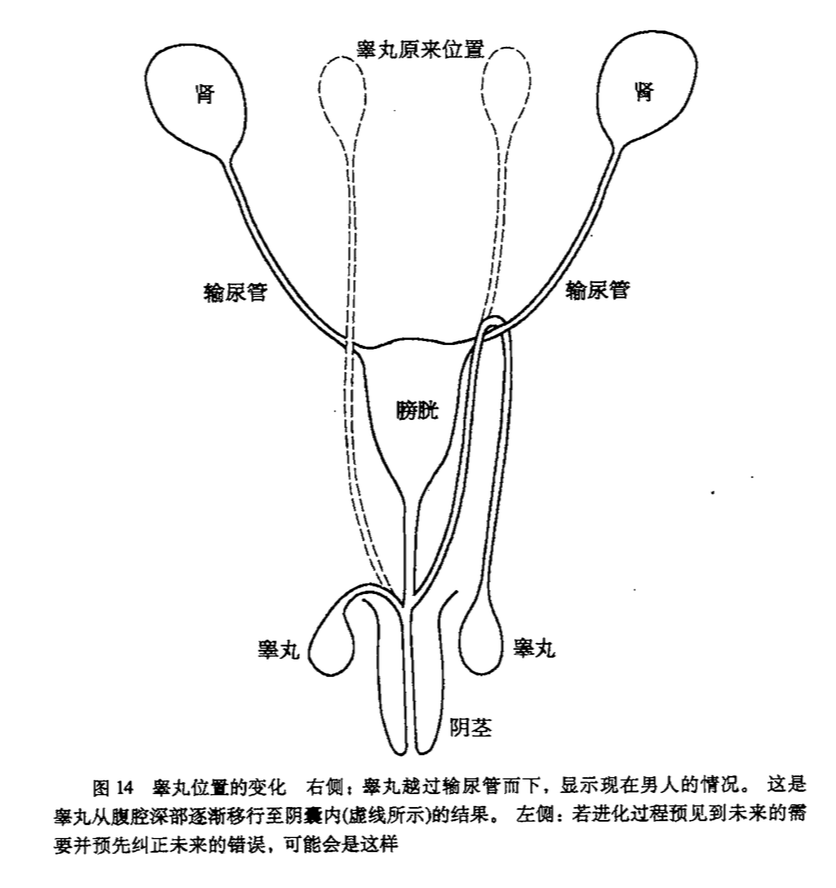
另一个类似的历史遗留问题是男性身上的输精管,直接挂在输尿管上了。
蝌蚪与打嗝
这是一个更奇妙的进化遗留问题。
不止人类会打嗝,很多哺乳动物也会(比如喵星人)。对一些人,吸入二氧化碳(通过在纸袋子里呼吸),或者伸展体壁(大口吸气然后屏住)可以止嗝。这个啥用没有的行为是怎么进化来的?尼尔·舒宾在 Your Inner Fish(PBS有同名纪录片)中对这问题进行了让人印象深刻的溯源。打嗝的特点是一次突然的吸气后紧接着一次声门的关闭。这个行为的适应性得一直追溯到蝌蚪身上。

下次打嗝的时候,就想想这是因为我们在与两栖类分道扬镳数亿年后体内依然有只阴魂不散的小小蝌蚪,一走神也许就不打了呢。
山魈与乳房

这是我之前在北京动物园拍的,休斯顿动物园的笼子比较大我小手机拍不清。山魈那花花绿绿的脸看着就不正常,花里胡哨的。这脸还得从它们的习性讲起。山魈是地球上最大的猴,性情凶猛,豹子来了都不太怕,所以它们不像其他猴子那样整天在树上跳来跳去紧张兮兮,而是大大方方的坐地上。(北京动物园这只我在那等了半小时,想看一眼它屁股,结果它就坐那吃吃吃!)小日子虽然安逸,却带来了一个问题,就是屁股不容易看见了,而猴子的发情信号重点就在屁股上。

最重要的发情信号被挡住了,作为弥补,它们在容易看到的地方进化出了新的信号。新信号要有效,最简单的办法当然是去模拟旧信号,于是,它们把屁股长到了脸上:


这一变化对解释人类身体特征是很有启发的。我们直立行走后屁股上的性信号也不容易看到了,于是也需要在更显眼的地方产生新的信号,而新信号也模拟了旧信号。

想庆幸我们至少没把屁股长到脸上?那就想想为什么我们在成年后还保有外翻的嘴唇吧(`3´)。至于这是对什么的拟态,我就真没法说太细了,只提供一个轶事证据,黑色人种因为肤色削弱了嘴唇的信号强度,于是他们厚嘴唇似乎也是最多的。更多内容请移步辉格的这篇文章。


大猿与精子战争
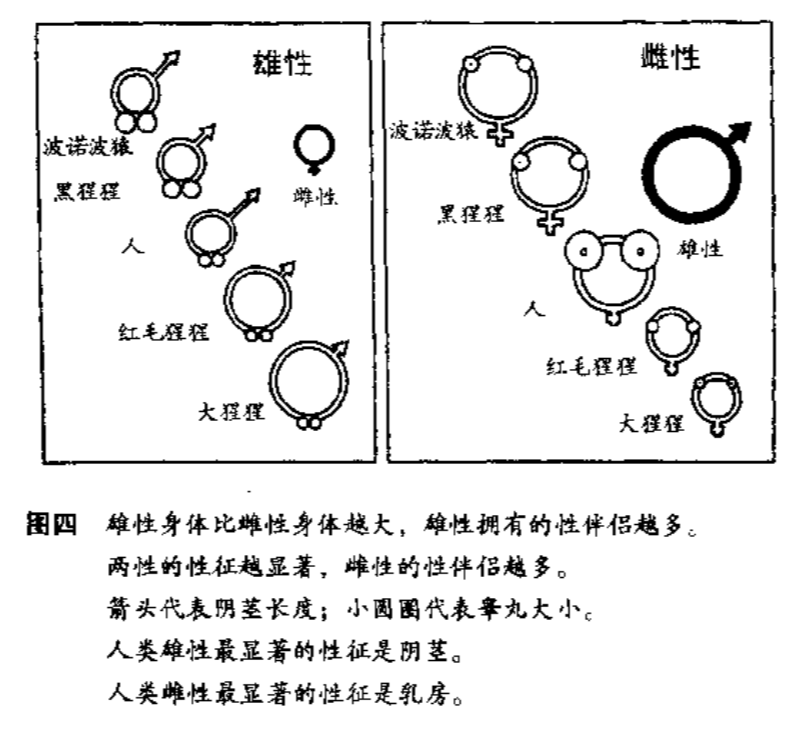
既然谈论了女性性特征,那也谈下男性的吧。跨物种比较一下:大猩猩一个群体里由一个主雄 (alpha male) 带着一群后宫和个别没交配权的雄性,不同群体分开住;人类是偶尔出轨的一夫一妻制;倭黑猩猩(下图的波诺波猿)是乱交的,性对它们来说几乎是社交润滑剂了,找到块新的香蕉地都要群P庆祝一下。结果三个物种里,倭黑猩猩具有最大的睾丸/体重比,大猩猩最小,人类居中。
图就不配了,去刘大可的这篇问答看吧。
蛇与图灵
如果提到图灵你想到的只是电脑那你对他的了解就远远不够。在自杀前两年图灵发表了一篇《形态发生的化学基础》(The Chemical Basis of Morphogenesis),这是他短暂生命中又一篇开创性的论文,是非平衡态热力学、非线性系统、混沌理论的先驱之作。他发现用简单的方程就能得出复杂的斑点条纹图案(有没有让你想起费曼?),而且与生物体身上的相似,这过程现在被命名为图灵斑图。具体来说他研究了反应扩散方程
(reaction–diffusion equation)
具有非线性项时的解其实我也不知道我在说什么。

图灵斑图的实际验证不多,在一些鱼和老鼠胚胎上有些成果。不过这些都太复杂,我们看点能懂的结论吧。大致来说,一个动物的身体长宽比较接近时,更可能是斑点,长宽差很多则更可能是条纹,而且条纹也更可能是垂直与体轴的方向,所以像蛇这种身体很长很窄的就更可能具有垂直体轴的条纹而不是其他。于是我今天在爬行两栖馆仔细看了下各种蛇啊蜥蜴的,好像是这么回事呢,不过例外也有。

另一个推论是你可以身体是条纹尾巴也是条纹,可以身体斑点尾巴也斑点,可以身体斑点尾巴条纹,但是不能身体条纹尾巴斑点。事实好像确实如此。

蚂蚁与自发秩序
这世界上除了人类之外还有一类动物,它们有农业(还会使用农药),有牧业(游牧、定牧都有),有奴隶,还造出了中央空调,请问这动物是?
某种猿?大象?海豚?鲸鱼?
都不是。你把那些拥有巨大大脑的动物列个遍也找不到答案,因为答案是脑子小的可怜的蚂蚁。它们的复杂程度你可以从 BBC 《灌丛下的生命》第五集了解个大概。
蚂蚁靠着那么点大脑却达到了大部分脑子更大的物种都没能实现复杂性,那是因为它们的做法很特别。就拿中央空调来说吧,人类造空调时是设计师脑子里先有一副蓝图然后把它投影到现实世界,而蚂蚁显然不会在它小小的脑子里塞进巨大蚁穴的蓝图,而是每只蚂蚁就靠着信息素短视的做着手边的事,最后一个复杂结构却涌现了出来,这就是哈耶克所说的“自发秩序”。人类当然也有自发秩序,而且比蚂蚁复杂得多。宗教、语言、市场、时尚……没有哪个有着中央设计者,事实上试图对它们进行设计的尝试都可悲的失败了。但就像蚂蚁没能力欣赏蚁穴的复杂一样,太多人对生活于其中的复杂秩序熟视无睹,反而孜孜不倦的追求着仁慈的父爱政府来操心他们的生活。蚂蚁也许能给他们上一课,不过我不抱太大指望。
当然蚂蚁的自发秩序和人类的还是有重大区别,对此辉格做过很详尽的分析了。讲到真社会性,还不能不提 E. O. Wilson 2010年在 Nature 上那篇引来无数撕逼的关于真社会性起源的文章,辉格对此也做过很出色的评述: 1, 2, 3。
动物行为与奎因难题
奎因提过一个关于语言的问题。当你看到一个语言不通的土著指着一只兔子说 "gavagai" 时,你怎么知道他的意思是 "Lo, a rabbit." 还是 "Lo, food"; "Let's go hunting"; "There will be a storm tonight" (these natives may be superstitious); "Lo, a momentary rabbit-stage"; "Lo, an undetached rabbit-part."?略做推广我们还可以得到各种不同学科的奎因难题。比如当动物行为学家看到两只动物纠缠在一起时,他怎么知道这行为是玩耍、示威、打架、认输、求爱还是纯粹偶然?
从常识出发我们确信我们大致能知道,不然不同语言的人就永远不可能交流了。但只有你想清楚这自然而然的过程并无任何神秘之处,从而意识到所有的“理解”都只能基于行为主义/操作主义来定义,而你则是在某物之前与之后的行为中找到联系。这个联系可能很复杂,所以你会建立一些模型与概念来帮助预测,比如“诚实”、“理性”、“自由意志”、“她喜欢我”……但这些概念不管看着多像在描述对方的精神活动,它们的来源永远都只能是可被观察的行为。
搞清楚了这一点我们就能避开一类让人很无语的问题了。每当谈及诸如“动物能否思考”这种问题时,老有人说“我们怎么知道动物能不能‘真正’地思考”,于是困在波普尔的第一世界连第一步都迈不出去。其实你是怎么知道坐你右前方那个萌妹子(归根到底她就是一坨有机物)能思考的,人类学家就是怎么知道另一坨有机物(比如一个 Yanomama 人)能不能思考的(不要把这视为理所当然,黑人就曾经被认为不具备完整的思考能力),动物学家就是怎么知道再另一坨有机物(比如一只黑猩猩)能不能思考的。
然后把“一坨有机物”换成“一堆电子元件”应该也能拯救那些纠结于“机器能不能真正思考”的家伙了吧。
其实受困于此类问题的人都还没理解“图灵测试”的深刻之处,这么基本的问题他老人家早解决了。
起太晚今天其实没逛完,所以可能待续。
下一篇 奥斯汀游记:图灵斑图再探